Introduction
This is perhaps one of the more controversial topics I have taught over the years. Since my first days in the chemistry department in Oregon, the professor whom I worked alongside and under during those 17 years (Al Levinson) and I would talk about this relationship that apparently existed between plant chemicals and where they existed in the Plant Kingdom and in turn where those members of the Plantae that bore these chemicals resided on the planet.

To both of us the first example that came to mind were the terpenoid resins of the Pinidae or Coniferae. The Conifers north of the equator produced a very distinct resin in terms of chemical make up and physical character which was quite different than those resins produced by many species distributed only in regions that are south of the equator. The old adage for these differences has been that this has something to do with the coriolis effect. The coriolis force dictates how wind patterns behave north and south of the equator. The following image from Wikipedia explains this and how it influences wind patterns:

The rotation of the earth places a force upon objects that causes them to behave slightly different. The effect of this force is to cause something to bend to the left or right in what it is doing, depending upon which hemisphere it is in–north or south. In chemistry, there are molecules that demonstrate this reflective or mirror image symmetry in several different ways. The most common mirror images that one learns about in basic chemistry are those noted in the following flowchart of these isomers:
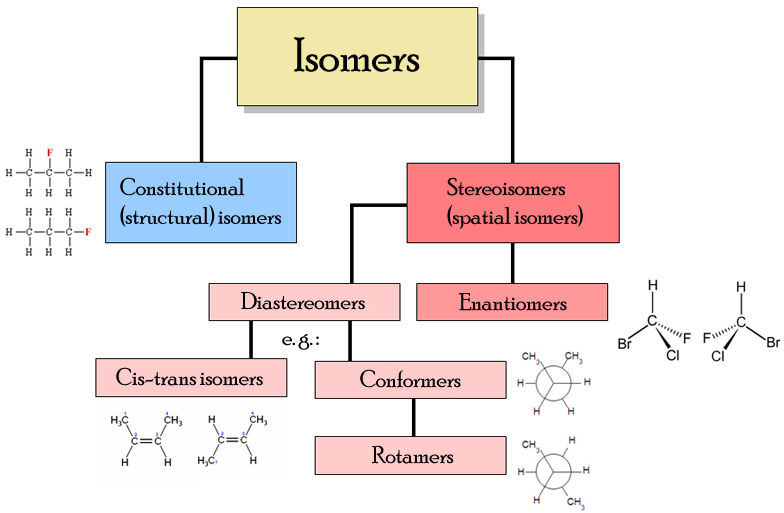
Wikipedia, “Stereoisomers”
The classical training tells us that there are these (+) and (-) relationships and distributions in some chemicals, like sugars, and that there are these l- or levo and d- or dextro (left and right forms) in others, cis- and trans- for still others, etc. Normally we might look at these differences and decide they are the result primarily of chance and circumstance, like flipping a coin and getting either heads or tails. In the chemical manufacturing setting that is an open vessel, there is in theory the equal likelihood that a chemical is going to bind at one end or the other of a chain or linear array, and if nothing else is involved with or interfering with this process, the chances for either selection will be equal between the two. But due to contacts that chemicals have to make with chemicals and surfaces in the cell, these events don’t usually occur using some random, free-floating process for the bonds to form. The chemical is not in an open vessel, but is in reality in some carefully laid out, architecturally planned, intracellular environment, for example to old enzyme-substrate system designed to engage in such activities.

Evolution has provided life forms with enzymes and a surface upon which this process of chemical reaction occurs. Therefore, any reaction that ensues has two factors limiting and controling its process–the enzymes engaged in the work and the surface upon which this work takes place, such as a protein molecule surface or a part of some organelle related membrance like the endoplasmic reticulum.

This “barrier” effect of living structures impacts how chemicals can be produced in many cases. In the case of terpenoid synthesis in Coniferae, if this process were to take place within a free-floating vessel environment, these physical limitations to how the final chemical was produced no longer hold. This means that the natural forces that regulate this process can control this process mostly through how much the enzymes floating around that cell are able to engage in this process. Terpene chains form only a specific way. With Coniferae there is somewhat of an order to how the polyterpenes are being produced, in that they are polymerized in linearly, rarely in a branching fashion. The only main variation here is that conifers north of the equator do this one way, and conifers south of the equator do this the totally opposite way.
The fact is what makes northern hemisphere Coniferae resins tacky and sticky and southern hemisphere Coniferae resins mostly brittle and crackly once they are allowed to dry. The hemiterpenes and monoterpenes of the southern species are more easily evaporated from the resin mass, leaving it in a drier form than northern hemispher pine pitch. Examples of this are the copal and dammar commonly used for artists’ paint base and formulas. This unique difference in the resins may be due to evolutionary mechanisms and the development of these paths in such a way that north-south differences did exist and did impact the chemical factory being developed. For this reason, many pharmacognosists in the past have claimed there to a a possible effect of the earth’s rotation on how the terpene building blocks are bound together in the different Coniferae families and orders. This is a unique explanation for the role of geography on the synthesis of chemicals in plants, and has relationships as well to the paleobotanical processes for chemical production in plants.
The more popular theories out there today for how plant chemicals evolved generally give energy-related features such as rotation and coriolis effect a lower role in this process. The most important elements or characters of nature and the natural settings that define chemical synthesis in plants in relation to geography are:
- Time–in terms of what stage in evolutionary development a plant is in relative to all other plants that exist for the time and place, and what stage in development its chemical pathways are in relative to its competitors and it precedents.
- The Pangaea versus Continents effect–Pangaeal one island, long-distance, topographic barriers effect versus Quarternary many island, natural oceanic barriers effect in the evolutionary procdess, relative to latitude featuers.
- Latitude–creates impacts through climate, wind flow patterns, temperatures, etc.
- Longitude–creates impacts only in terms of how continents or smaller physical surfaces of the world are isolated and made into island communities for specific chemical groups
- Environmental water content–impacts of water chemistry (saline versus fresh, versus thermosalinic, etc.) on plant type and needs; impacts of water availability and contact on plant chemical reactions intracellularly, interstitially, and morphologically
- Elevation–impact of elevation on average temperates seasonally and climatically; impact of elevation on precipitation behaviors and patterns, impact of elevation on atmospheric content
- Small-scale or immediately local physiography–impact of slope and aspect on water and solar exposure, climate, weather patterns, aridity, etc.
- Climate–impoact of standard climate features on plant chemical synthesis behaviors
- Weather–impact of specific weather patterns, especially seasonal, on the plant chemical sysnthesis behaviors
- Continent–in terms of large scale island barriers and temporal relationships
- Impact of all of the above on local environment, and whether or not specific selective features are developed thereby creating evolution-based stress patterns of specific species
- Impact of all of the above on local zoogeography and ecological relationships
Each of these in turn are somewhat regulated by the nature of the end products–for example, if an end product such as a terpene or fixed seed oil is very unstable in the given setting, then this chemical will either be non-beneficial and require too much energy for synthesis to occur and be maintained throughout the organism’s lifespan, or it will result in selection of that species and its chemical paths in such a way as to either cause a change in that pathway or result in extinction of the species engaged in this method of chemical production.
Roger Good’s Geography of Plants
I often start this topic that I like to teach with some of the evidence already provided about plant evolution, and plant distribution globally and paleontologically, by Ronald Good in his The Geography of Flowering Plants (New York: London, Green and Co., 1947). By the time this work was published, the pangaeal hypothesis of the earth’s surface was well underway to being accepted. Good used this to develop his interpretations of how particular classes, orders, families and genera of plants were distributed around the world. As noted in his forward, his work was largely based in turn on previous plant evolution and geography writings produced by S. A. Cain’s Foundations of Plant Geography (New York, 1944) and E. V. Wulff ‘s An Introduction to Historical Plant Geography (translated by E. Brissenden, Waltham, Mass, 1943). Earlier writings that helped researchers like Good along in this research process were [in chronological order]:
- E. Goeze. Pflanzengeographie. Stuttgarte, 1882
- H.C.Watson. Topographical Botany. 2ed. London, 1883.
- G.S. Boulger. Plant Geography (The Temple Primers). London, 1912.
- L. Diels. Pflanzengeographie (Sammlung Goschen 389) Auf. 2, Berlin, 1918
- A. Hansen. Die Pflanzendecke der Erde. Leipzig, 1920.
- H. N. Ridley. The Distribution of Plants. Annals of Botany, 37, 1923.
- M.E. Hardy. The Geography of Plants. Oxford, 1925.
- D.H. Campbell. Outline of Plant Geography. London, 1926.
- A. Hayek. Allgemeine Pflanzengeographie. Berlin, 1926.
- J. Hutchinson. The Families of Flowering Plants. London 1926, 1934
- H. Walter. Allgemeine Pflanzengeographie Deutschlands. Lena, 1927.
- P. Graebner. Lehrbuch der allgemeinen Pflanzengeographie. Auf. 2, Leipzig, 1929
- E. F. Wulff. “Essay of Dividing the World into Phytogeographic regions . . . ” Bull. Appl. Bot. Genetics and Plant-breeding, Supp. 52. Leningrad, 1932.
- J.C. Willis. The course of evolution by differentiation or divergent mutation rather than by selection. Cambridge, 1940.
- D.H. Campbell. “Continental Drift and Plant Distribution.” Science, 95, 1942.
- Sir T. H. Holland. The Theory of Continental Drift. Proc. Linn. Soc. London, 1942-3.
Much of the above writers work is based on the combination of paleontological evidence with contemporary plant ecology knowledge. G. Madaus’s Deutsche Medizinische Wochenschrift (H. 26. 27, Leipzig, 1938), made an attempt to correlate this with medically important species. The numerous regional studies out there during this time sometimes engaged in much the same sorts of speculation.
The most direct relationships between palnt evolutionary traits or characters and time and place are seen in work on climate patterns, particularly in the writings of O. E. White:
- O.E. White. “Geographical Distribution and cold-resisting characters . . .” Brooklyn Botanical Garden Rec.. 15, 1926.
- O.E. White. “Mutation, Adaptation to Temperature Differences, and Geographical Distribution in Plants.” Verhand des V. Intern. Kongr. fur Vererbungswissenschaft, Berlin, 1927.
The different philosophies that are out there about plant evolution spatially and temporally also help to set the stage for trying to better understand phytochemical evolution. By breaking the seemingly endless realtionships that plants can have with the earth as a whole, relative to time and stages in surface topographical development,, we can come up with a large number of evolutionary, environmental, ecological, and developmental reasons for why and how certain chemicals evolved in plants. In some cases, climate plays the major role in the selection process for a specific group of chemicals. In other cases, the presence of new animal herbivores defines the natural selection-survival process. In yet other cases, local land features and the impacts of these on local foliage, local herbivores and local temperatures nad soil features all play a role in what chemicals a particular plan t will develop. A collection of unusual examples of these environmetnal features that seem to impact plant chemical evolution include the following:
- continental shift
- volcanism
- helioclimatic influences (sunspot cycles and long term thermal-radiation changes)
- natural disaster history (asteroid showers, comets, etc.)
- other reasons for changes in topography, elevation features, climate, local weather and such
- the production of salinic, cyanic or sulphur-rich water
- population density changes through natural selection processes (increase competitors, increased pathogens, increased plant-consumers)
The following are examples of these features and how they impact phytochemical evolution:
- Hamamelidae and the development of uniquely large fixed oils due to cold climates
- Umbelliferae genera with temperate and mediterranean-arid climate terpenoid production, with less evaporative products developed in hot, arid climate settings
- High seed-producing, highly competitive Caryophyllidae members capable of utilizing flavonoids for photosynthesis instead of chlorophyll during periods of droughts and low nutrition
- Asteraceae-Lactucoideae and the conversion of aromatics into bitter, saponifying agents to prevent slug eating habits
- The development of latices (latex, pl.) to protect a plant against bark-borers
- Specific plants and the development of unique terpenoids required by insects for the development of their pheromones used in mating
- Quercaceae and the development of specific fungal resistant tannins in the protective layers of their bark
- Orchids and the development of unique aromatics specific to certain insecta
Not all chemicals in plants have this distinct evolutionary link to their development in terms of long-term events and changes. Most phytochemicals are developed in order to best meet the needs for the time, which is usually a very narrow period in the world different periods of development. Nevertheless, there are features that do stand out as helpful examples for demonstrating how and why some types of chemicals form or begin to exist along certain phytogenetic (allelic) lines. It is the more broadly applicable rules of evolution and their relationships to local geography that are being reviewed, followed by any possible ecologic rules that might exist as well for the species and environmental conditions at hand. The point of this section is to look at spatially-defined or spatially-limited features that impact phytochemical development, and to use these to understand how and why such chemicals exist in plants.
Plans for executing this review
The following types of chemical evolution can be related to this, some of which can be detailed separately (for those who took my classes, this is that use of the plant chart I simplified and handed out on the distributions of seeds oils by chemical class, such as the C12 to C24 evolution, C=C bonding behaviors related to latitude and Temp., and role of vinylization processes, etc.):
- The tropical Palmidae
- The tropical to temperate climate transition in the evolution of Magnoliidae
- Mixed tropical-temperate-subarctic Pinidae features and requirements
- Seasonal winds and the graminae
- Fixed Seed Oils and the impacts of climate, temperature and latitude on fixed oil types; fixed seed oils as an ecological development
- Benzylisoquinolines (biq), Magnoliidae, and the differentiation and increased complexity of the biq pathway during the lignose to herbaceous and northerization (northern migration/shift) of latitude climate evolutionary stages
- Terpenoids (non-oxygenated, and oxygenated separately) as ecological and environmental features; primitive versus advanced coadaptation and ecological impact features for each
- The early evolution of hallucinogens and their non-hallucinogenic, alterative successors
- Complex Alkaloids (evolution and products from merged alkaloid-non-alkaloid pathways)
- Complex tertiary-quaternary chemical evolution development–why certain species in unrelated taxa produce the send end product with specific human ecological adaptation related uses (the evolution of, cancer, autoimmune and antiviral drugs)
- Examples of emergent evolutionary features between very different taxa lineages
- etc.
.

Leave a comment